La regulación del genoma en los eucariotas es mucho más compleja que el de los procariotas. La bacteria (procariota) Escherichia coli alberga un único cromosoma circular con 4.600 pares de bases, que almacena la información para la síntesis de aproximadamente 2.000 proteínas. A título comparativo, uno de los organismos eucariotas más simples (Sacharomyces cerevisiae) contiene 16 cromosomas, con entre 200 y 2.220 pares de bases cada uno. En conjunto, el genoma de la levadura del pan (Sacharomyces cerevisiae) tiene 17.000 pares de bases, que codifican la síntesis de alrededor de 6.000 proteínas.
El genoma de una célula humana contiene 46 cromosomas (23 pares). El ADN de cada cromosoma consta entre 50.000 y 250.000 pares de bases. En conjunto, el genoma humano contiene aproximadamente 3 millones de pares de bases, organizados en unos 40.000 genes.
La regulación génica es compleja, no solo porque cada célula debe expresar determinados genes, y reprimir la expresión de otros, sino porque en las células eucariotas, a diferencia de lo que sucede en las procariotas, los procesos de transcripción y traducción no están acoplados.
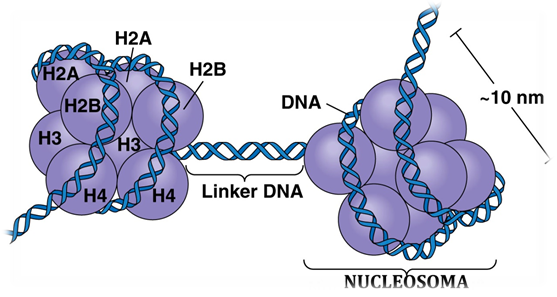
El ADN en las células eucariotas se halla asociado a proteínas denominadas histonas. El concepto de cromosoma abarca, pues, el ácido desoxirribonucleico (ADN) y las proteínas (histonas) a las que se halla unido. Las histonas representan prácticamente la mitad del tamaño del cromosoma. La cromatina es el conjunto de cromosomas de una célula.
Las histonas son proteínas con carga positiva neta, debido al elevado número de restos aminoácidos de arginina y lisina. Existen cinco clases de histonas: H2A, H2B, H3, H4 y H1. Las cuatro primeras (H2A, H2B, H3 y H4) se asocian por pares formando un octámero, mientras la histona H1 actúa de puente (linker) entre octámeros contiguos. Esta estructura fue desentrañada en el año 1974 por Roger D. Kornberg, Premio Nobel de Química en el año 2006 por sus estudios sobre las bases moleculares de la transcripción eucariota. El conjunto de 146 pares de bases de ADN (200 según otros textos bioquímicos) y el octámero formado por 1 par de cada una de las histonas H2A, H2B, H3 y H4 definen un nucleosoma. De este modo, la cromatina tiene el aspecto de un collar de cuentas. Cada cuenta del collar (nucleosoma) tiene un diámetro aproximado de 100Å (Å, de Amstrong equivalente a 10-10 metros). La histona restante (H1) actúa de puente (linker) entre nucleosomas contiguos.
Las 8 histonas forman a manera de una rampa superhelicoidal levógira con el ADN en rededor. Recuérdese que el ADN es una molécula con carga negativa neta, mientras las histonas tienen carga positiva. De este modo se consigue la estabilización eléctrica del nucleosoma.
El ADN forma una superhélice levógira a medida que se enrolla alrededor del perímetro del octámero de histonas (véase dibujo al comienzo del artículo). El ADN es una doble hélice enrollada sobre sí misma.
La importancia de las histonas se infiere de la conservación de sus secuencias de aminoácidos a lo largo de la evolución. Una de las funciones de los nucleosomas es el empaquetamiento del ADN en el minúsculo espacio disponible en el núcleo celular. Si se estira el ADN de un nuclesosoma, se desenrollará de manera incompleta, requisito imprescindible para la replicación y transcripción. La duplicación del ADN durante la mitosis tiene un mecanismo diferente.
En las levaduras (los organismos eucariotas más simples) la activación de los genes necesarios para la utilización de la galactosa como nutriente requiere la unión de una proteína denominada GAL4 que reconoce una secuencia específica del ADN. Este fragmento de ADN tiene dos secuencias 5’-CGG-3’ separadas por 11 pares de bases. En el genoma de la levadura existen aproximadamente 4.000 secuencias a las que podría engarzarse la proteína GAL4. De éstos, solo 10 regulan los genes necesarios para el metabolismo de la galactosa. ¿En qué secuencias, de las posibles, se une la proteína GAL4? Mediante una técnica denominada ChIP (Chromatin ImmunoPrecipitation) se descubrió que solo 10 de los 4.000 posibles sitios de unión, están ocupados por la proteína GAL4 cuando la levadura se mantiene en un medio rico en galactosa. Así, mientras en la cromatina de los procariotas todos los sitios de unión son accesibles, en las células eucariotas la estructura de la cromatina enmascara (protege) a determinadas secuencias, mientras deja libre acceso a otras idénticas.
Se infiere de éste, y otros experimentos, que la estructura de la cromatina de los genes activos e inactivos es diferente. Las histonas parecen ser responsables de estas diferencias.
Además, determinadas proteínas se unen a lugares específicos del ADN. Estos lugares se definen como intensificadores.
INTENSIFICADORES EN LA TRANSCRIPCIÓN
La función de los intensificadores (fragmentos de ADN) consiste en servir como lugares de unión a determinadas proteínas reguladoras. Un intensificador (fragmento de ADN) precisa de un regulador (una proteína) para su función, aun cuando miles de pares de bases separen a ambos.
Las propiedades de los intensificadores se han descifrado a partir de los estudios del intensificador que controla la isoforma muscular de la enzima creatina-quinasa. Existe un intensificador localizado entre los pares de bases 1.350 y 1.050 anteriores al lugar de iniciación del gen que codifica esta enzima.
En las células eucariotas cada factor de transcripción (regulador, intensificador, ARN polimerasa, y otros) incorporan otras proteínas para formar grandes complejos que interaccionan con la maquinaria de transcripción. El resultado es la activación o represión de la expresión génica. A la participación de muchas proteínas se le denomina control combinatorio.
Recomendamos también la lectura de las siguientes páginas web 0dentro de www.info-farmacia.com:
- Histonas.
- Regulación genética de los esteroides.
Zaragoza a 287 de septiembre de 2018
Dr. José Manuel López Tricas
Farmacéutico especialista Farmacia Hospitalaria
Farmacia Las Fuentes
Florentino Ballesteros, 11-13
50002 Zaragoza